


| name | Amanita brunnescens var. straminea |
| name status | insufficiently known |
| author | E.-J. Gilbert |
| english name | "American Citrin Amanita" |
| images |
, George Res., Pinckney, Washtenaw Co., Michigan, U.S.A.") 1. Amanita brunnescens f. straminea, holotype collection (MICH), George Res., Pinckney, Washtenaw Co., Michigan, U.S.A. , George Res., Pinckney, Washtenaw Co., Michigan, U.S.A.") 2. Amanita brunnescens f. straminea, holotype collection (MICH), George Res., Pinckney, Washtenaw Co., Michigan, U.S.A. |
| intro |
The name Amanita brunnescens var.
straminea has been largely forgotten.
This is "good" and "bad." One might consider
that it is inappropriately confusing to use this name
for a taxon that is clearly much more like the
similarly eastern North American "A.
citrina lavendula than it is
like A.
brunnescens. It is important to know what this citrina-like taxon is. Is it distinct from the European A. mappa? Is it distinct from the American A. lavendula? The answers to these questions are currently being pursued. It seems quite possible that the present name would apply to one of the "cryptic" taxa now being identified among the citrinoid taxa of eastern North America. See, for example, A. americitrina. |
| cap | The cap of this taxon is 40–80 mm wide, Light Chalcedony Yellow overall, convex, becoming planar in age, and viscid when young. There is no information on the cap's flesh. The margin of the cap is not striate and usually somewhat decurved. Volval remnants on the cap are Pale Olive Buff or more yellowish warts and patches, with thicker patches more tannish; or the volva may be present as a thin, very variable coating in young material—submembranous in some cases, but floccose-fibrillose in others. These remnants are usually rather evenly distributed or leave large areas uncovered in older material. Small remnants often remain near the margin even in older material. |
| gills | Annotation of the type by Dr. A. H. Smith indicates that the gills touched the stem. They also were crowded (125–133 per cap) and ivory-yellow. In the dried material of the type, the short gills are plentiful, of diverse lengths, and cut off with a rounded corner or more or less slowly narrowing toward the stem. |
| stem | The stem of this mushroom is 60–100 × 10–15 mm and the same color as the gills. It bears a thin coating of fine fibrils above skirt-like ring. Below the ring, the stem's surface is sparsely fibrillose or bears fibrillose scales or patches. No information is available about the stem's flesh. The stem's bulb is 20–35 mm wide, white, and spongy. The stem's ring is almost as greenish yellow as pileus, with ragged edges. The volva is present as a submembranous, lobed, membrane standing erect and distinctly separated from the stem by as much as 5± mm. The lobes are sometimes large and irregular and are always placed along the distinct bulb margin. |
| odor/taste | The odor of the present taxon is reported to be "faint, but pungent" and "somewhat reminiscent of chlorine." No taste was recorded. |
| spores | The spores measure (6.0-) 7.5 - 9.0 (-11.0) × (5.8-) 6.8 - 8.7 (-10.2) μm and are globose to subglobose (infrequently broadly ellipsoid) and inamyloid. There are no clamp connections at the bases of basidia. |
| discussion |
Amanita brunnescens f. straminea was originally described from Michigan, U.S.A. While its range is not yet clearly understood, this taxon appears to be restricted to the eastern half of North America. The relationship between this taxon and Amanita lavendula is not yet fully understood.—R. E. Tulloss |
| brief editors | RET |
| name | Amanita brunnescens var. straminea | ||||||||
| author | E.-J. Gilbert. 1941. Iconogr. Mycol. (Milan) 27 suppl. (2-3): 338, tab. 46. | ||||||||
| name status | insufficiently known | ||||||||
| english name | "American Citrin Amanita" | ||||||||
| synonyms |
≡Amanitina brunnescens f. straminea E.-J. Gilbert. nom. nud 1940. Iconogr. Mycol. (Milan) 27 suppl. (1): 78, tab. LVIII (fig. 6).
non Amanita citrina f. straminea (Scop.) E.-J. Gilbert (q.v.)
non Amanita straminea Secr. (q.v.)
non Amanita straminea Cleland (q.v.). [Note: a species of A. sect. Lepidella.] The editors of this site owe a great debt to Dr. Cornelis Bas whose famous cigar box files of Amanita nomenclatural information gathered over three or more decades were made available to RET for computerization and make up the lion's share of the nomenclatural information presented on this site. | ||||||||
| etymology | stramineus, "straw-colored" | ||||||||
| MycoBank nos. | 345944 | ||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||
| lectotypes | MICH (implicit) | ||||||||
| lectotypifications | proposed herein | ||||||||
| type studies | Tulloss (here) | ||||||||
| intro |
Because it is now generally assumed that Gilbert’s
personal herbarium was lost after his
death, the holotype of A. brunnescens var.
straminea, which was in that herbarium must
also be assumed lost. Fortunately, A. H.
Smith, who collected the holotype, retained an
isotype with annotations and photographs in
MICH. This isotype is proposed as the
lectotype of the taxon herein. The
macroscopic description that accompanies this type
study is based on A. H. Smith’s original notes and
on photographs of the lectotype
collection. The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material not directly from the protolog of the present taxon and not cited as the work of Dr. Z. L. Yang or another researcher is based upon original research by R. E. Tulloss. | ||||||||
| pileus | 40–80 mm wide, Light Chalcedony Yellow overall, convex to obtuse, becoming planar in age, viscid when young; context not recorded; margin nonstriate, usually somewhat decurved; universal veil Pale Olive Buff or more yellowish, with thicker patches near Avellaneous, as thin, very variable coating in young material, submembranous in some cases, floccose-fibrillose in others (and then breaking up into [radially striate, shingle]-like patches especially near margin), [minutely verruculose under 10× lens in exsiccata] with remnants usually rather evenly distributed or with large areas becoming glabrous with age, with small remnants at margin often remaining in older material. | ||||||||
| peridium | double click in markup mode to edit. | ||||||||
| lamellae |
touching stipe, crowded (125–133 per pileus), Ivory Yellow, 5–7 mm broad, "broadest in front and outer ends, obtusely rounded behind," with edges crenulate under lens, [with edge minutely fimbriate and pallid under 10× lens in exsiccata]; lamellulae [in exsiccata, plentiful, of diverse lengths, rounded truncate to subattenuate to attenuate]. [Note: RET finds the phrase in quotation marks difficult to interpret. Suggestions would be appreciated.] | ||||||||
| stipe | 60–100 × 10–15 mm, concolorous with lamellae, with thin coating of fine fibrils above partial veil (in age this layer sometimes breaking up into concentric [irregular zones, subcylindric or narrowing upward slightly, flaring very little at apex,] below partial veil sparsely fibrillose or with fibrillose scales or patches; context not recorded; bulb 20–35 mm wide, white, spongy; partial veil almost as greenish yellow as pileus, with ragged edges; universal veil as submembranous, lobed, limbate volva standing erect and distinctly separated from stipe by as much as 5± mm, sometimes with lobes large and irregular, with lobes placed along distinct bulb margin. | ||||||||
| odor/taste | Odor faint, but pungent, somewhat reminiscent of chlorine. Taste not recorded. | ||||||||
| macrochemical tests |
none recorded. | ||||||||
| pileipellis | 265–270 µm thick, slightly gelatinized just at surface, uniformly yellowish in 3% KOH soln.; filamentous undifferentiated hyphae 2.4–8.4 µm wide, branching, dominantly subradially arranged, densely interwoven in vertical section, some with walls more yellow than in others; vascular hyphae not observed. | ||||||||
| pileus context | filamentous undifferentiated hyphae 1.8–12.5 µm wide, with walls thin to 0.8 µm thick, branching, sometimes with fusiform intercalary cells (up to 14 µm wide and occasionally branched), in fascicles or singly, with fascicles and individual hyphae interwoven in loose matrix; acrophysalides elongate (e.g., 77 × 17 µm) or narrowly clavate (e.g., 186 × 35 µm), with walls thin or up to 0.7 µm thick; vascular hyphae not observed. | ||||||||
| lamella trama |
bilateral; wcs = 55–70 µm; filamentous undifferentiated hyphae, with many intercalary segments slightly inflated, 2.0–22 µm wide; some inflated cells apparently terminal, similar to intercalary cells (e.g., 89 × 19.2 µm or 101 × 22 µm), with slightly thickened walls, with shallow angle of divergence eventually reach angle of up to 80° to central stratum, sometimes penetrating up to 10± µm into subhymenium. [Note: Notes on the type study (which date from 1993) do not differentiate the hyphae and intercalary cells in the central stratum from those in the subhymenial base. RET feels that distinction should be drawn.] | ||||||||
| subhymenium | bilateral; wst-near = 50–65 µm; wst-far = 60–85 µm; pseudoparenchymatous (cellular), overall 28–42 µm thick, comprising 3–5 layers of inflated cells (globose to subglobose, to ovoid to clavate, up to 22 × 18.0 µm); clamps not observed. | ||||||||
| basidia | 27–52 × 8.8–11.5 µm, dominantly 4-, infrequently 2- or 3-sterigmate; clamps not observed. | ||||||||
| universal veil | On pileus: pale grayish brown in mass; filamentous undifferentiated hyphae 2.5–8.2 µm wide, branching, in fascicles or singly, with walls thin to slightly thickened, plentiful; inflated cells dominating, terminal, often in chains of three or more, easily dissociated, collapsing, thin-walled or with walls up to 0.8 µm thick, subglobose to broadly ellipsoid (e.g., 48–56 × 42–50 µm) to broadly fusiform (e.g., 60 × 31 µm), colorless to slightly sordid; vascular hyphae not observed; clamps not observed. At stipe base: slightly orangish brown in mass, as on pileus, but with larger percentage of hyphae. | ||||||||
| stipe context | longitudinally acrophysalidic; filamentous undifferentiated hyphae 3.0–9.0 µm wide, thin-walled, branching, plentiful; acrophysalides up to 262 × 43 µm, thin-walled, dominant away from stipe surfaces; vascular hyphae not observed; clamps not observed. | ||||||||
| partial veil | filamentous undifferentiated hyphae 2.5–11.5 µm wide, subradially arranged, sometimes coiling loosely, with thin or slightly thickened, frequently branching, occasionally with yellowish walls, sometimes with refractive guttules decorating wall inner surfaces, densely interwoven but easily separated, in fascicles or singly; inflated cells, narrowly clavate or clavate or broadly clavate, terminal singly, common to plentiful, with thin walls, 26–40 × 8.5–19.5 µm; vascular hyphae not observed; clamps not observed. | ||||||||
| lamella edge tissue | not described. | ||||||||
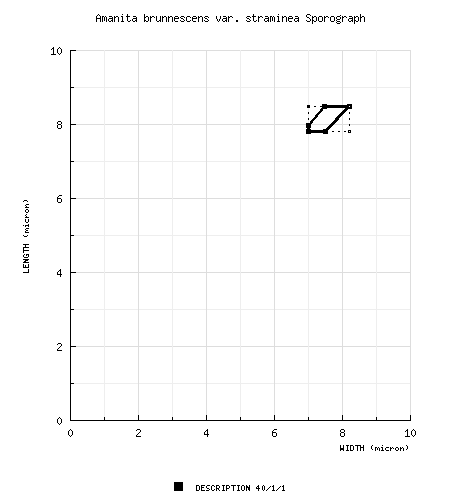
| basidiospores | from type study of RET: [40/1/1] (7.5–) 7.8 – 8.5 (–9.2) × (6.8–) 7.0 – 8.2 (–9.0) µm, (L = 8.2 µm; W = 7.6; Q = (1.03–) 1.04 – 1.14 (–1.15); Q = 1.07), hyaline, colorless, smooth, thin-walled, globose to subglobose, infrequently broadly ellipsoid, adaxially flattened; apiculus proportionately small, cylindric, sublateral; contents mono- to multiguttulate, with additional small granules; color in deposit not recorded. | ||||||||
| ecology |
no information recorded non-type, RET: Solitary to subgregarious. New Jersey: Ca. sea level. In deep sandy soil of Atlantic coastal plain in Pinus-Quercus barrens. Tennessee: In moist loam and litter of upland Quercus-Pinus forest. | ||||||||
| material examined | RET: U. S. A.: MICHIGAN—Washtenaw Co. - Pinckney, George Reserve, 24.ix.1938 H. V. & A. H. Smith 11066 (isotype, MICH 4953; isotype fragment, RET 683-8). WEST VIRGINIA—Tucker Co. - Fernow Exp. For., 13.ix.1992 R. P. Bhatt & A. Kumar FMR-5 (FWVA), 25.ix.1992 S. L. Stephenson, R. P. Bhatt & A. Kumar PW-2 (FWVA); Fernow Exp. For., Watershed 4, 8.x.1993 S. L. Stephenson 93-09 (FWVA). | ||||||||
| discussion |
The epithet "straminea" is not available in
Amanita at species rank. Therefore
I chose to propose a new name (Amanita
americitrina) that would (in part) conserve the
name that North Americans have been using for the
present species. Gilbert felt that A. brunnescens var. straminea was distinguishable from A. mappa on several counts including habit, pileus color, spore size, form of the universal veil, etc.; but felt that neither taxon was sufficiently well described so that the details could be laid out. ??more—comparisons, etc.?? A sporograph comparing the present taxon with A. mappa (green figure) follows. Smith notes that the limbate volva of the present species is "not membranous as in [A.] verna." The major problem in the taxonomy of the present taxon is the issue of whether it should be segregated from A. lavendula. A comparison sporograph follows: Preliminary genetic studies support the idea that we have had for some time—that the present taxon should be treated at the rank of species and is distinct from A. lavendula as the above sporograph suggests. These same studies indicate that the present taxon probably will turn lavender if the weather is sufficiently cool. To the best of our current knowledge, we think that the present taxon can be identified with A. americitrina. We await further developments following a year of intensive collecting in eastern North America by a group of volunteer collectors to whom we owe a great deal of thanks. | ||||||||
| citations | —R. E. Tulloss | ||||||||
| editors | RET | ||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.